

Marine birds
More than 70 bird species regularly utilize Puget Sound during some or all stages of their life histories, but only a portion of these are actively being investigated.
. Photo by Finley and Bohlman, courtesy U.S. Fish and Wildlife Service")
Background
Puget Sound is important for nesting, wintering, and migration of numerous bird species associated with the marine environment. More than 70 bird species regularly utilize Puget Sound during some or all stages of their life histories (Gaydos and Pearson 2011), but only a portion of these are actively being investigated. Studies have focused primarily on abundance and distribution, habitat utilization, foraging behavior, and contamination levels.
Multispecies comparisons
Information pertaining to marine bird distribution and abundance prior to the 1970s resides primarily in anecdotal accounts (Rathbun 1915, Jewett 1953) and systematic surveys held during Christmas Bird Counts (CBCs), which became consistent and widespread in the 1960s. Since the 1970s, the most comprehensive census of marine birds in northern Puget Sound was conducted as part of the Marine Ecosystems Analysis (MESA) program of 1978-1979 (Wahl 1981). The MESA study was a large-scale survey jointly funded by the Department of Commerce (DOC) and the Environmental Protection Agency (EPA) as a response to oil spill threats in the Strait of Juan de Fuca. It included aerial, land-based, and ferry-based transect surveys north of Admiralty Inlet, within portions of the Straits of Juan de Fuca and Georgia, and the Canadian Gulf Islands. Notably, the study included only the southernmost portion of the Strait of Georgia and not Puget Sound itself.
Beginning in 1992, the Puget Sound Ambient Monitoring Program (PSAMP) began collecting observations of marine birds in the non-breeding season; this currently is the only source of continuous multi-species monitoring of marine birds in Puget Sound. The annual surveys consist of aerial transects covering nearshore habitat and offshore habitat/open waters throughout Puget Sound and the southern shore of the Strait of Juan de Fuca (Figure 1). Aircraft-based observers record all bird species seen below the high tide line, but monitoring goals and data summaries emphasize certain alcid, diving duck, loon, and grebe species.
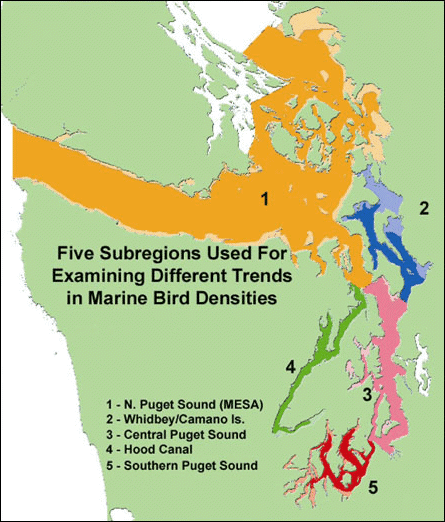
Figure 1. Map of PSAMP subregions (Reprinted from Evenson et al. 2010 with permission from Washington Department of Fish and Wildlife)
Nysewander et al. (2005) evaluated long-term changes in abundance in several species of marine birds by comparing the PSAMP results from 54 aerial transects with results from nearly identical MESA transects. Results of this analysis revealed significant declines in 13 of the 20 species or species groups studied, including declines in at least one species from each marine bird family found in northwestern Washington. For some species, such as the western grebe (Aechmophorus occidentalis) and long-tailed duck (Clangula hyemalis), declines were as high as 95% and 91%, respectively. Although methodologies used in MESA and PSAMP surveys were relatively comparable, differences did exist, for example in the locations and habitat types surveyed by the aerial methods, and in the proportion of the MESA baseline data that was from aerial, land-based and ferry-based surveys. Furthermore, the PSAMP used aerial surveys, but the potential bias associated with avoidance of aircraft by birds has not been evaluated.
Results from Nysewander et al. (2005) and other studies (e.g. Wahl 2002) sparked concern over declines in marine birds in Puget Sound. In acknowledgment of these concerns and the multiple problems associated with comparing results across disparate survey methodologies, the Western Washington University (WWU) or WWU/MESA comparison study was initiated (Bower 2009). The WWU/MESA comparison study replicated land-based and ferry-based transect portions of the MESA surveys over two winters (2003-2004 and 2004-2005). Results of the WWU/MESA comparison of data were largely consistent with the MESA/PSAMP comparison (Nysewander et al. (2005), although a few results diverged. To perform a third comparison of marine bird observations over time, Bower (2009) analyzed annual Christmas Bird Count (CBC) data from 11 count circles north of Puget Sound (1975-1984 and 1998-2007). Table 1 summarizes characteristics of the data sets used by to make comparisons (Bower (2009).
Table 1. Attributes of bird surveys compared in Bower (2009)
| Study | Year(s) | Geographic area | Methods |
| Marine Ecosystems Analysis (MESA) | Jan-Dec 1978-1979 | Admiralty Inlet (S), Tsawwassen-Schwartz Bay BC Ferry (N), Neah Bay (W), and WA mainland (E) | Shore-based point counts, ferry and small boat transects, aerial transects |
| Puget Sound Ambient Monitoring Program (PSAMP) | Winter 1992-1999 | Straight coastline between Admiralty Inlet (S), Strait of Georgia (N), Neah Bay (W), and WA mainland (E) | Aerial transects compared with 1970s MESA aerial transects |
| Western Washington University (WWU) | Sept-May 2003-2004 and Sept-May 2004-2005 | Admiralty Inlet (S), Tsawwassen-Schwartz Bay BC Ferry (N), Sand Juan Islands (W), WA mainland (E) | Shore-based point counts and ferry transects compared with 1970s MESA shore-based point counts and ferry transects |
| Christmas Bird Count (CBC) | 1975-1984 and 1998-2007 | Salish Sea, including 8 BC and 3 WA CBC circles | Standard CBC methods for 11 CBC circles, data from 1975-1984 with data from 1998-2007 |
Bower (2009) reported a 29% decline in the total number of marine birds in inland waters of the Salish Sea – which includes areas and data from outside the Puget Sound Basin – between 1978/79 and 2003-2005 (Figure 2). It should be noted, however, that this overall decline can be substantially influenced by changes exhibited by certain individual species. For example, of the 37 most common overwintering marine species, 14 showed significant declines and six showed significant increases. Notably, the largest declines were observed among taxonomically diverse groups, including common murre (Uria aalge) (–92.4%), western grebe (–81.3%), surf scoter (Melanitta perspicillata) (–59.8%) and brant (Branta benicla) (–73.2%). Species that showed increases in abundance included double-crested cormorant (Phalacrocorax auritus) (+97.7%) and pigeon guillemot (Cepphus columba) (+108.9%). Results from the CBC data comparison revealed significant declines in seven of the 37 most common species or species groups, with significant increases in three species (Bower 2009).
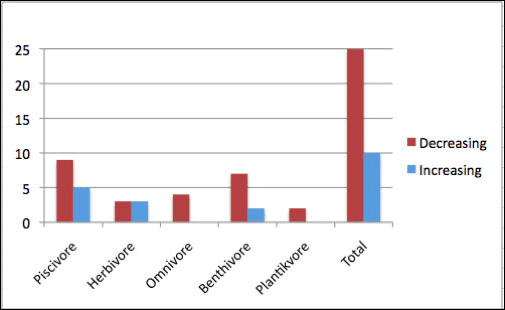
Figure 2. Changes in mean abundance among feeding guilds for 35 common overwintering marine birds in the Salish Sea between 1978/79 and 2003-2005 (data from Bower 2009)
Seventeen species or species groups were common to all three studies (the WWU/MESA comparison, the PSAMP/MESA comparison, and the CBC data comparison (Bower 2009). The PSAMP/MESA comparison revealed declines in more species (14 of 17) than did either the WWU/MESA comparison (six of 17) or the CBC comparison (three of 17) (Table 2). The PSAMP/MESA comparison showed no change or an increase in just three of 17 species or species groups, whereas the WWU/MESA comparison found no change or an increase in six of 17 and the CBC comparison found no change or an increase in eight of 17 species or species groups. Despite these differences, several consistencies emerge. First, the number of species declining exceeded the number of species increasing in all three comparisons. Second, three taxa -- western grebe, all scaup, and marbled murrelet-- showed declines across all three studies. And finally, only two species (Harlequin Duck, Bald Eagle) showed significant increases across all the three comparisons (Bower 2009).
Table 2. Comparison of percent change detected in three studies of non-breeding marine bird abundance for 17 species or species groups in Puget Sound (Bower 2009)
Studies | |||
Species | PSAMP/ MESA | WWU/ MESA | Recent/ historic CBCs |
| Common Loon | -64a | +49a | +13 |
| All loons | -79a | -33 | -47 |
| Red-necked Grebe | -89a | -46a | -35 |
| Horned Grebe | -82a | -72a | -30 |
| Western Grebe | -95a | -81a | -86a |
| Double-crested Cormorant | -62a | +98a | +171a |
| All cormorants | -53a | -8.3a | -25 |
| Great Blue Heron | -19 | +51 | -16 |
| Brant | -66a | -73 | +1027a |
| All scaup | -72a | -65a | -51a |
| Harlequin Duck | +189a | +20 | +7 |
| Long-tailed Duck | -91a | -44 | +49 |
| All scoters | -57a | -33a | -8 |
| Bufflehead | +20 | -11 | +5 |
| Bald Eagle | +35 | +187a | +28 |
| Pigeon Guillemot | -55a | +109a | +15 |
| Marbled Murrelet | -96a | -71a | -69a |
a Statistically significant
In summary, widespread changes in the abundance of marine birds during the non-breeding season have occurred over the last 30 years in the Salish Sea (Nysewander et al. 2005, Bower 2009). Causes of these declines are not adequately known.
Scoters
Puget Sound supports some of the largest wintering scoter populations on the west coast of North America (Wahl 1981), where they feed on regionally-abundant bivalves and forage fish roe. Puget Sound is also one of the three most important staging areas and one of two major molting areas for other west coast scoter populations, including scoters that winter in California, Mexico, and British Columbia. Scoter populations in Puget Sound, including the wintering, staging, and molting populations, consist primarily of surf scoters and white-winged scoters (M. fusca). Black scoters (M. perspicillata) are also present, but in much smaller numbers. Surf scoters are one of the most abundant diving ducks in Puget Sound between September and May, with the highest densities in southern and central Puget Sound (Nysewander et al. 2005). Washington’s wintering scoters spend eight to 10 months in marine waters, with males spending approximately a month longer than females, before migrating to the Canadian interior to breed on freshwater lakes.
Scoters in Puget Sound are found most often in shallow coastal waters (< 20 meters depth) over a broad range of substrates, including pebbles, sand, mud, cobble, and rock. Previously thought to subsist on a relatively narrow diet of bivalves, scoters are now understood to adjust foraging patterns and locations to take advantage of ephemeral food sources. During much of the winter, they forage on newly-settled mussels and soft substrates inhabited by clams and other shellfish. In spring, some scoters in the region feed on herring eggs where available and flocks of surf scoters regularly track the northward progression of spawning events to consume this abundant and energy-rich source of food (Vermeer 1981). Anderson et al. (2008) found that prey such as crustaceans and polychaetes associated with eelgrass habitats comprise a substantial part of scoter diets in late summer.
. Photo copyright Scott Streit, used with permission.")
Scoters observed in both nearshore and offshore waters during PSAMP winter monitoring efforts between 1992 and 2008 ranged in mean overall densities from 9.2 to 19.4 birds per km2 (Evenson et al. 2010). The density indices reported for nearshore areas, which scoters favor, ranged from 34.8 to 70.4 birds per km2. Figure 3 shows scoter densities between 1992 and 2008. Of all scoters counted along transects sampled during 1992-2008 winter surveys, between 33% and 90% were identified to species in any single year. Of these, surf scoters comprised 55-82%, white-winged scoters comprised 17-40%, and black scoters made up 0.2-9%. WDFW currently is conducting species/age/sex ratio surveys by boat to provide a better estimate of species proportions (Evenson 2010).
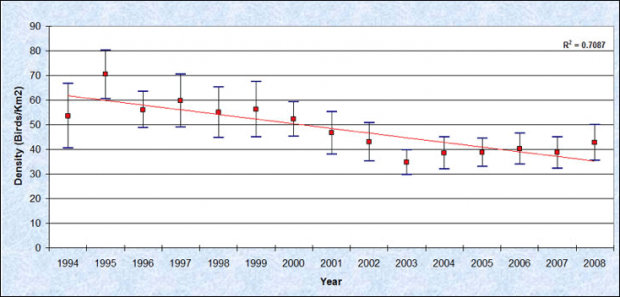
Figure 3. Mean winter densities of scoters in nearshore (<20 m) habitats of the inner marine waters of Washington state, 1993-2008 (Reprinted from Evenson et al. 2010 with permission from Washington Department of Fish and Wildlife)
Bower (2009) demonstrated that as a group scoters showed significant declines in both the PSAMP/MESA (-57%) and WWU/MESA (-33%) comparative studies. Surf scoters declined by 60% in the WWU/MESA comparison; however, nearly half of this decline is attributed to the collapse of the Cherry Point herring stock that occurred between the two survey periods (Stout 2001, Bower 2009). The evidence for this decline is compelling: more than 40,000 surf scoters were observed by MESA researchers in 1978 and less than 1,000 surf scoters were seen by WWU researchers at the same location in 2004 and 2005.
Comparisons of annual changes in density in the inner marine waters of Washington between 1992 and 2008 suggest that the scoters declined from the early 1990s through 2003, but that since 2003, densities have been relatively stable (Figure 3)(Evenson et al. 2010). However, spatial variation in rates of decline exist, for example in the Whidbey/Camano (North Puget Sound)area, where scoter densities have continued to decline (Figure 4)(Evenson et al. 2010). In 1993, the densities in the Whidbey/Camano area were the highest in the inner marine waters of Washington, but by 2008 densities in the Whidbey/Camano were the lowest (Figure 4)(Evenson et al. 2010).
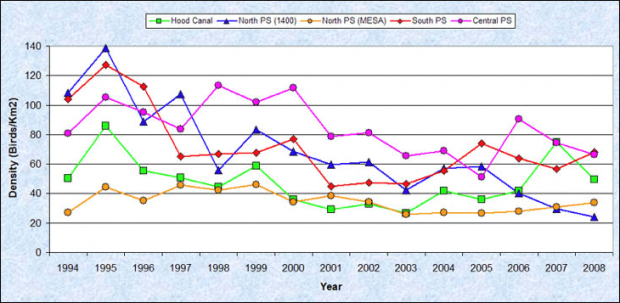
Figure 4. Comparison of winter scoter densities by region in the nearshore (<20 m) inner marine waters of Washington state (Puget Sound), 1993-2008 (Reprinted from Evenson et al. 2010 with permission from Washington Department of Fish and Wildlife)
Loons and Grebes
. Photo by Art Weber, U.S. Fish and Wildlife Service.")
Several species of loons and grebes spend a substantial portion of the winter in Puget Sound where they utilize a variety of marine habitats. Loon species include the common loon (Gavia immer), Pacific loon (G. pacifica), and red-throated loon (G. stellata). Common loons are widespread and fairly common during winter in almost all nearshore marine habitats, and in most freshwater habitats, except rivers, typically occurring as single birds or in small numbers. They are rare breeders in Washington waters with the majority nesting throughout Canada and Alaska. Common loons were listed as sensitive by WDFW because they are a rare breeding species in the state and are vulnerable to a number of threats, including destruction or alteration of nesting habitat, poor water quality (i.e., degradation of lakes), and human activity (Richardson et al. 2000). Pacific loons are also widespread and common during winter, but occur further offshore than common loons and are more likely to congregate. Flocks of Pacific loons feed on schools of small fish near banks, tidal rips, and other hydrographic features of deeper waters. This species breeds in eastern Siberia and from northern coastal Alaska across to Baffin Island and Hudson Bay in North America. Red-throated loons are widespread and fairly common during winter in Puget Sound; they breed throughout Alaska, Canada, Greenland, and northern Europe Asia, with the very southern portion of their range extending south to southern Vancouver Island. Although red-throated loons can frequent many different types of marine waters, they tend to favor estuaries and shallow offshore areas, aggregating at times in areas where prey species are concentrated by tidal conditions.
. Photo by Tim Bowman, U.S. Fish and Wildlife Service.")
Western grebes utilize marine and fresh waters in Puget Sound between October and April and tend to occur in groups. The primary wintering habitat for the larger flocks of western grebes are in offshore (>20m depth) marine waters with minimal tidal current flow, where they prey on schooling forage fish, although they may also occur in many saltwater situations and on inland lakes. Western grebes gather in large resting groups during the daytime hours and then disperse at night to forage. Major concentration areas have been identified through PSAMP aerial surveys (Evenson et al. 2010). Western grebes breed from southern British Columbia and the prairie Provinces in Canada south to Mexico.
. Image by Finley and Bohlman, courtesy U.S. Fish and Wildlife Service.")
Comparisons of survey data (Nysewander et al. 2005, Bower 2009) reveal that Puget Sound loon and grebe species have declined significantly in recent decades. Bower (2009) detected declines in loons as a group in all three comparative studies as follows: 64% decline in MESA/PSAMP comparison; 33% decline in WWU/MESA comparison; and 47% decline in historic/recent CBC comparison (Table 2). Records from the annual PSAMP winter aerial surveys from 1992 to 2008 show that loons constituted 0.8% of all marine birds surveyed (Evenson et al. 2010). The majority of loons were identified to species (common loon [28%], Pacific loon [27.9%], and red-throated loon [32.5%]) and occurred in both nearshore and offshore waters.
Among the three loon species, densities were lowest in the common loon, ranging from 0.17 to 0.57 birds per km2. A comparative analysis of common loon densities reported in MESA (1978/79) and PSAMP (1992-1999) surveys showed a 64% decline (Nysewander et al. 2005). Conversely, Bower (2009) reported 49% and 13% increases as shown by WWU/MESA and the historic/recent CBC data comparisons, respectively, which include survey data through the mid- 2000s. It is unclear whether these changes reflect some degree of recovery since 1999, shifts in distribution, or are an artifact of differing or variously effective methodologies (see Uncertainties section, below).
Densities of Pacific loons observed during PSAMP winter surveys ranged from 0.26 to 1.21 birds per km2 in 1994-2008, with higher densities (10 and 89 birds per km2) observed in areas where flocks concentrate. Pacific loon winter densities appeared to be relatively stable over the period 1994-2008, although this result may be confounded by other loon species (Evenson et al. 2010). A comparison between MESA and PSAMP data was not made for Pacific loons due to the difficulty of distinguishing Pacific loons from red-throated loons in aerial surveys. Analysis of PSAMP subregional density indices suggest that Pacific loons favor certain subregions, such as northern Puget Sound, Whidbey/Camano Islands, and Central Puget Sound near Bainbridge Island (Evenson et al. 2010).
. Photo by Dave Menke, U.S. Fish and Wildlife Service.")
Red-throated loon densities of 0.17 to 1.20 birds per km2 were observed during PSAMP winter surveys of nearshore and offshore areas between 1994-2008 (Evenson et al. 2010). Densities appear to have been relatively stable over the past two decades (Evenson et al. 2010), although this species is not clearly separated from other loon species in some survey data.
Grebes
All grebe species wintering in Washington marine waters have exhibited some degree of decline over the last two decades, but western grebes have declined most sharply (Evenson et al. 2010). Overall densities for western grebes, combined for both nearshore and offshore waters, ranged from 3.9 to 13.2 birds per km2, while densities in the vicinities of the flocks ranged from 50 to 1,343 birds per km2 (Figure 5). A comparative analysis of western grebe densities reported by MESA (1978/79) and PSAMP (1992-1999) surveys showed a 95% decline (Nysewander et al. 2005). Bower (2009) noted that declines were observed in all three comparative studies (Table 2), across of the Salish Sea, and in every month of the MESA/WWU comparative surveys. Density indices reported by PSAMP winter monitoring surveys between 1992-2008 suggest that this species is still declining (Evenson et al. 2010).
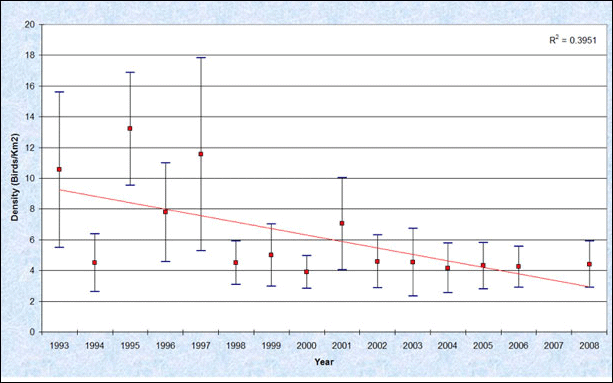
Figure 5. Winter Trends in Western Grebe Densities in the Inner Marine Waters of Washington State, 1993-2008 (Reprinted from Evenson et al. 2010 with permission from Washington Department of Fish and Wildlife)
Alcids
. Photo by Laura Whitehouse, U.S. Fish and Wildlife Service.")
Several alcid species utilize marine waters of Puget Sound during winter months, with some species breeding along coastlines and on islands. Pigeon guillemots (Cepphus columba) are common and widespread residents that feed in nearshore habitats, along rocky shorelines, passes, banks, areas with tidal currents and rips, as well as in shallow embayments. These birds are seldom seen in flocks, except near colonies during breeding, although they can aggregate in productive feeding areas such as tidal convergences and passes. Pigeon guillemot nest in nearly every small-island or saltwater coastline habitat, with larger colonies are found in San Juan, Jefferson, Island, and Clallam Counties. Smaller colonies and single pairs are found throughout Puget Sound, making them the second most common breeding seabird in Puget Sound.
. Photo by Scott Streit, used with permission.")
Rhinoceros auklets (Cerorhinca monocerata) are found throughout Puget Sound in both coastal habitats and far from land. Most often they often feed close to shore, especially where tidal currents near islands create localized upwelling and trophic intensification. Flocks may overnight in protected bays and forage farther out to sea during the day. Rhinoceros auklets in Washington nest at three main sites: Destruction Island, Protection Island, and Smith Island. Smaller numbers nest at a few other sites in Puget Sound.
. Photo by Gus Van Vliet, U.S. Fish and Wildlife Service.")
Marbled murrelets (Brachyramphus marmoratus) are small, fast-flying seabirds present year round in coastal areas throughout Washington. They are non-colonial alcids that breed in mature inland forests up to 84 km from marine shorelines that support prey such as small schooling fish or invertebrates in shallow waters (Raphael 2006). Areas of winter concentration in Washington include the southern and eastern end of the Strait of Juan de Fuca, Sequim, Discovery and Chuckanut Bays, and the San Juan Archipelago. In 1992, the Pacific coast population of marbled murrelets south of the Canadian border was listed as Threatened by both USFWS and the State of Washington. Critical habitat in Washington, Oregon and California was designated in 1996. Primary threats to marbled murrelets include the loss and modification of nesting habitat, primarily due to commercial timber harvesting of older forests, effects resulting from oil spill pollution, and to a much lesser degree, risks associated with capture in commercial fisheries gear (Ralph et al. 1995).
In 2003, a WDFW survey of pigeon guillemot colonies in Puget Sound reported at least 471 colonies, representing approximately 16,000 breeding birds (Evenson et al. 2003). Long-term changes in pigeon guillemot populations are not known due to absence of historical data. Records from annual PSAMP aerial surveys show that pigeon guillemot densities were highest in nearshore habitats (<20m depth), where they ranged from 0.26 to 1.18 birds per km2 in 1992-2008 (Evenson et al. 2010). Densities from the inner marine waters of Washington during winters 1993-2008 increased from 1993-1997, and then remained stable through 2008. A comparative analysis of pigeon guillemot densities recorded by MESA (1978/79) and PSAMP (1992-1999) surveys showed a 56% decline over that period (Nysewander et al. 2005). However, Bower (2009) reported a 109% increase in pigeon guillemot density based on the WWU/MESA comparative study, which covered a slightly longer time period (Table 2). The inconsistencies likely reflect differences in sampling between the studies (Bower 2009) and a the lack of knowledge of pigeon guillemot post-breeding dispersal patterns (Evenson et al. 2010).
Rhinoceros auklet breeding populations in Puget Sound are concentrated on Protection Island and Smith Island. Protection Island hosts 70% of the breeding birds within Washington’s inner marine waters (Speich et al. 1989). Estimates of the breeding population of Rhinoceros auklets on Protection Island have shown a 30% decline in breeding pairs with more than 17,000 breeding pairs in 1975 (Wilson and Manuwal 1986) decreasing to approximately 12,000 pairs in 2000 (Wilson 2005).
In 2006, marbled murrelet population size was estimated to be about 22,000 in Washington, Oregon, and California (Huff et al. 2006), compared with approximately 860,000 in Alaska and 55,000 to 78,000 in British Columbia in 2004 (McShane 2004). At-sea counts of marbled murrelets using boat-based observer transects were conducted from 2000 to 2009 as part of effectiveness monitoring of the Northwest Forest Plan. In 2009, USFWS conducted a five-year status review of the Northwest Forest Plan and determined that marbled murrelets in Puget Sound had continued to decline significantly since the previous review conducted in 2002 (Pearson et al. 2010). The population estimate for marbled murrelets in all zones in the Northwest Forest Plan area (Washington, Oregon and California) was 17,791 (95% confidence interval: 14,631 – 20,952). Estimates from the 9 years of monitoring have ranged from 17,354 to 23,673. The 2009 population estimate for Puget Sound and Juan de Fuca Strait east of Cape Flattery from at-sea surveys was 5,623 birds (95% confidence interval: 3,922 – 8,352 birds). The annual rate of decline for the 2001-2009 period was 7.0% (standard error = 1.8%; Pearson et al. 2010). For Washington State overall, there was a significant decline in murrelet density for the 2001-2009 period (Pearson et al. 2010). The largest concentrations of birds occurred in northern Puget Sound and the Strait of Juan de Fuca.
High Arctic Black Brant
. Photo by Peter Mickelson, U.S. Fish and Wildlife Service.")
High arctic black brant are a subpopulation of brant geese that utilize Puget Sound shallow bays and saltwater marshes from late November through May. They breed in the high arctic of western Canada, primarily on Melville Island and Prince Patrick Island, and then stage for over a month at Izembek Lagoon in Alaska before heading to wintering grounds in Puget Sound. Brant wintering habitats are usually characterized by an abundance of eelgrass and marine algae (e.g., Padilla, Samish and Fidalgo Bays in Skagit County). Large concentrations of brant may gather at Dungeness Spit and Willapa Bay, but smaller flocks are present in the southern Puget Sound. Because of their strong dependence on certain plants, fidelity to wintering and breeding locations, and because some populations live in harsh environments, brant are more vulnerable to periodic breeding failures and occasional heavy losses from starvation than are most other geese (Reed et al. 1998).
Results from the comparative MESA and PSAMP studies showed that brant abundance varied widely over spatial and temporal scales (Bower 2009). Brant exhibited declines in the PSAMP/MESA comparision (-66%) and WWU/MESA comparison (-73.2%), but increased by more than 1000% in the CBC comparison data (Bower 2009). The large decline in the WWU/MESA comparison was principally driven by a decline in numbers on the primary wintering grounds of Padilla and Samish Bay. Outside these two locations, brant numbers showed a slight increase. CBC comparison data showed increases in brant in British Columbia, possibly indicating a change in the wintering location of brant.
Great Blue Heron
. Photo by George Gentry, U.S. Fish and Wildlife Service.")
In Puget Sound, great blue herons (Ardea herodias) belong to a non-migratory and marine-oriented subspecies (A. herodias subsp. fannini) that ranges from Alaska to southern Washington state, with the largest concentration occurring in northwestern Washington and southwest British Columbia (Butler 1997). During the non-breeding season, great blue herons are widely dispersed in Puget Sound, utilizing coastal and lowland areas for foraging and roosting. They are often found as solitary individuals. In contrast, between late winter and summer, herons occur in high densities centered on nesting colonies and associated foraging sites. Herons forage in a variety of habitat types depending on local conditions, tides, and season. Saltwater and freshwater marshes provide year-round foraging opportunities of fish, crustaceans, amphibians and reptiles, though terrestrial habitats also provide small mammals in heron diets (Eissinger 2007).
Marine shoreline and intertidal areas are important to the success of coastal heron colonies. In 2004, WDFW performed an aerial survey to determine foraging habitat, distribution, and concentration areas of great blue herons in Puget Sound (Hayes 2006). Based on this survey it was estimated that 73% of the active heron colonies in Puget Sound are directly associated with marine and estuarine intertidal habitats for foraging activities during the breeding season. In particular, the reproductive success of colonies is dependent on prey associated with eelgrass habitats (Eissinger 2007), such as Drayton Harbor, Port Susan, and Samish, Padilla, and Skagit bays.
Few records of historical trends exist for the great blue heron in Puget Sound. Methods for monitoring heron colonies in British Columbia and Puget Sound have recently been developed, although they are not yet standardized between the two areas. In western Washington, colony status has been assessed approximately every four years by WDFW biologists, and larger colonies in certain locations are monitored by independent investigators or conservation groups. Eissinger (2007) conducted a review of available population data and concluded that since the mid-1990s, the population of northwestern great blue herons has been stable, with the current estimate at 4,700 nesting pairs or 9,400 breeding herons in 2003-2004 (Figure 6). This breeding population represents 121 colonies located on Vancouver Island, the British Columbia mainland, and in the Georgia Strait and Puget Sound basins. Notably, approximately 66% of the total population is concentrated in only 16 colonies, and 35% of the total breeding population belongs to five mega-colonies supporting 200-600 breeding pairs each. In the past decade, the Puget Sound population has seen a substantial transformation from a diffuse distribution of smaller colonies across the landscape to larger colonies in upland marine areas. Reasons for this shift are unknown, but possible causal factors include combinations of increased predation by expanding bald eagle population, human disturbance and encroachment on habitat, degradation and fragmentation of nearshore and coastal habitats by development and land use activities, pollution, changes in prey abundance or distribution, and other systemic changes related to ecosystem decline (Eissinger 2007).
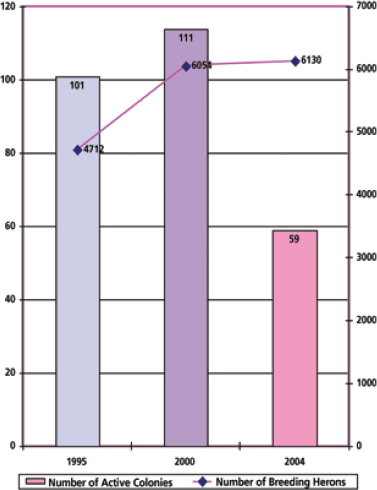
Figure 6. Great blue heron population trends in Puget Sound (reprinted from Eissinger 2007 with permission from the Puget Sound Nearshore Ecosystem Restoration Project and Washington Department of Fish and Wildlife)
Uncertainties
With the recovery of Bald Eagle populations, anecdotal information indicates predation pressure (direct and indirect) has increased at Great Blue Heron colonies. The effect of increasing Bald Eagle presence on colony persistence or productivity by Great Blue Herons is not known.
Trends in waterbird abundance derived from Christmas Bird Counts must be assessed to evaluate whether correction factors that account for observer effort (e.g. party hours) are appropriate. Correction factors applied where they are not necessary could result in a conclusion that abundance had decreased when in fact it had not changed.
Many marine birds migrate, overwinter or breed in regions quire distant from the area(s) they use in Puget Sound. The degree to which potentially significant limiting factors in those areas influence observed changes in abundance in Puget Sound is largely unknown.
Additional work is needed to determine whether changes in abundance of particular marine birds reflect actual population changes or shifts in regional distribution that would locally mimic population declines.
Summary
Multiple species of marine bird that overwinter in Puget Sound have shown sharp declines in abundance over the past two decades. Declining species outnumber increasing species, declines occur across diverse taxonomic groups and feeding guilds, and declines of up to 95% have been reported. Reasons for these declines are not well established and may include factors operating locally, along migration flyways, and at the breeding grounds. Habitat loss and changes in food availability or abundance may have contributed to population changes.
Literature Cited
Anderson, E. M., J. R. Lovvorn, and M. T. Wilson. 2008. Reevaluating Marine Diets of Surf and White-Winged Scoters: Interspecific Differences and the Importance of Soft-Bodied Prey. The Condor 110:285-295.
Bower, J. L. 2009. Changes in marine bird abundance in the Salish Sea: 1975 to 2007. Marine Ornithology 37:9-17.
Butler, R. W. 1997. The great blue heron : a natural history and ecology of a seashore sentinel. UBC Press, Vancouver, BC.
Eissinger, A. M. 2007. Great blue herons in Puget Sound. Puget Sound Nearshore Partnership, Olympia, WA.
Evenson, J. R. 2010. Analysis of marine bird data, Puget Sound Assessment and Monitoring Program. Seattle, WA.
Evenson, J.R, D.R. Nysewander, B.L. Murphie, and T.A. Cyra. 2010. Marine Bird and Mammal Component of the Puget Sound Ambient Monitoring Program web reports. Available at: http://wdfw.wa.gov/mapping/psamp/index.html. Washington Department of Fish and Wildlife, Olympia, WA.
Evenson, J. R., D. R. Nysewander, M. Mahaffy, B. L. Murphie, and T. A. Cyra. 2003. Status, abundance, and colony distribution of breeding pigeon guillemots (Cepphus columba) from the inland marine waters of Washington State, as documented by PSAMP efforts, 2000-2002 in Proceedings from Georgia Basin/Puget Sound Research Conference.
Gaydos, J. and Pearson, S. 2011. Birds and Mammals that Depend on the Salish Sea: A Compilation. Northwestern Naturalist 92(2), 79-94.
Hayes, G. 2006. Puget Sound Great Blue Heron Foraging Aerial Survey 2004. Washington Department of Fish and Wildlife, Olympia, WA.
Jewett, S. G. 1953. Birds of Washington State. University of Washington Press, Seattle.
McShane, C. 2004. Evaluation report for the 5-year status review of the marbled murrelet in Washington, Oregon, and California. EDAW, Inc., Seattle, WA.
Nysewander, D., J. Evenson, B. Murphie, and T. Cyra. 2005. Report of marine bird and mammal component, Puget Sound Ambient Monitoring Program, for July 1992 to December 1999 period. Washington Department of Fish and Wildlife, Olympia, WA.
Ralph, C. J., G. L. Hunt, Jr., M. G. Raphael, and J. F. Piatt. 1995. Ecology and conservation of the marbled murrelet. U.S. Dept. of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, CA.
Raphael, M. G. 2006. Conservation of the Marbled Murrelet under the Northwest Forest Plan. Conservation Biology 20:297-305.
Rathbun, S. F. 1915. List of Water and Shore Birds of the Puget Sound Region in the Vicinity of Seattle. The Auk 32:459-465.
Reed, A., D. H. Ward, D. V. Derksen, and J. S. Sedinger. 1998. Brant (Branta bernicla): Birds of North of America.in A. Poole, editor. The Birds of North America Online. Cornell Lab of Ornithology, Ithaca, NY.
Richardson, S. A., D. Hays, R. Spencer, and J. Stofel. 2000. Washington State status report for the common loon. Washington Dept. of Fish and Wildlife, Olympia, WA.
Speich, S. M., T. R. Wahl, and J. F. Watson. 1989. Catalog of Washington seabird colonies. U.S. Dept. of the Interior, Fish and Wildlife Service, Research and Development, Washington, D.C.
Stout, H. A. 2001. Status review of Pacific herring (Clupea pallasi) in Puget Sound, Washington. U.S. Dept. of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Northwest Fisheries Science Center, Seattle, WA.
Vermeer, K. 1981. Food and populations of Surf Scoters in British Columbia. Wildfowl 32:107-116.
Wahl, T. R. 1981. Marine bird populations of the Strait of Juan de Fuca, Strait of Georgia, and adjacent waters in 1978 and 1979. National Oceanic and Atmospheric Administration; United States Environmental Protection Agency, Research and Development, Seattle, WA; Washington, D.C.; Springfield, VA.
Wahl, T. R. 2002. Trends in numbers of marine birds wintering on Bellingham Bay. Washington Birds 8:29-40.
Wilson, U. W. 2005. The Effect of the 1997-1998 El Nino on Rhinoceros Auklets on Protection Island, Washington. Condor 107:462-468.
Wilson, U. W., and D. A. Manuwal. 1986. Breeding Biology of the Rhinoceros Auklet in Washington. Condor 88:143-155.









