

Photo courtesy of Dave Cowles, Walla Walla University.
Background
Pinto abalone (Haliotis kamtschatkana) were once widely distributed throughout the waters of British Columbia and Washington state. In recent decades, populations have undergone sharp declines, likely in response to the combined stressors of overharvest, poaching, and sub-optimal environmental conditions (Campell 2000). Known for their large, muscular foot and their pearlescent oval shell, pinto abalone are slow-growing, long-lived marine snails and are typically found in nearshore rocky habitats in semi-exposed or exposed coastal regions. More than 60 abalone species are found worldwide but the pinto, or northern, abalone is the only species found in Washington State, where they range from Admiralty Inlet to the San Juan Islands and the Strait of Juan de Fuca and are typically found at depths to about 20 m (Bouma 2007).
Abalone are important herbivores in nearshore habitats, feeding primarily on drift macroalgae such as kelp and benthic diatom films. They can structure subtidal communities through the maintenance of substrata dominated by crustose coralline algae and through the facilitation of conspecific settlement. The larvae are planktonic and settle after approximately 7 -10 days in response to cues from both crustose coralline algae and from adults. Juvenile pinto abalone are cryptic until they reach a shell length of >50 mm.
Abalone are broadcast spawners. Consequently, the number and proximity of spawning adults determines the likelihood of successful fertilization (e.g., Babcock and Keesing 1999, Miner et al. 2006). At low population numbers, fertilization success may be low or nil, potentially limiting population recovery from overharvesting (Rothaus et al. 2008).
Status
The Washington Department of Fish and Wildlife (WDFW) regularly monitors the abundance of pinto abalone at 10 index stations throughout the San Juan Archipelago (Rothaus et al. 2008) (Figure 1). Because pinto abalone are highly patchy, cryptic and frequently associate with microhabitats such as rock crevices or patches of coralline algae that may themselves be patchily distributed, total abundances are not measured (Rothaus et al. 2008). Rather, repeated surveys at a system of index sites are conducted so that temporal trends in abalone abundance may be detected. The WDFW sites are composed mostly of bedrock and boulders encrusted with coralline algae, and support assemblages of kelp and other macroalgae (Rothaus et al. 2008). The sites range in size from 135 m2 to 380 m2, and individual animals are counted and measured during each survey.
Data from surveys made in 2006 showed an overall mean abalone density of 0.04 m-2 (Rothaus et al. 2008), which is well below the minimum densities for successful reproduction (0.15 individuals m-2 ) and recruitment (1 individual m-2 ) reported respectively by Babcock and Keesing (1999) and by Miner et al. (2006) for congeners of the pinto abalone.
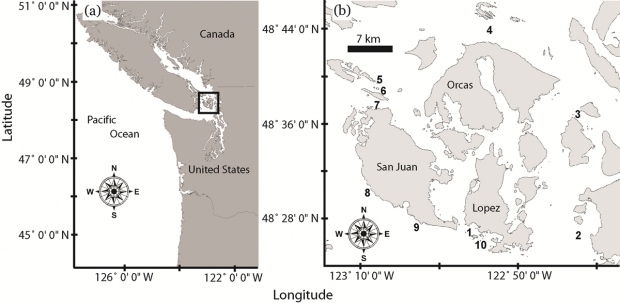
Figure 1. Map of WDFW Haliotis kamtschatkana index stations established in 1992 in the San Juan Archipelago, Washington State (Figure produced for Rothaus et al. 2008, CJFAS and used with permission).
Trends
The decline of pinto abalone in Washington State has been of concern since the early 1990s (Rothaus et al. 2008). While commercial harvest of abalone has never been permitted in the state, the sport fishery may have extracted as many as 38, 200 individuals per year in the San Juan Archipelago (Bargmann 1984). It is therefore possible that abalone densities may have already been too low for successful fertilization or recruitment at the time of the sport fishery closure in 1994. WDFW listed the pinto abalone as a candidate species for protection in 1998 and NOAA Fisheries listed it as a federal species of concern in 2004. In 2008, WDFW identified pinto abalone as a Species of Greatest Conservation Need. In British Columbia, Canada, pinto abalone were uplisted to endangered in 2009, where populations are generally found at higher densities than Washington stocks (COSEWIC 2009).
The WDFW index site surveys in the San Juan Archipelago were repeated in 1994, 1996, 2003, 2004, 2005, 2006 and 2009. These surveys indicate a decline in abalone abundance of 83% from 1992 to 2009 (WDFW)(methods according to Rothaus et al. 2008)( Figure 2). Rothaus et al. (2008) also found an increase in mean shell length of 10.4 mm between 1992 and 2006, indicating a substantial shift in the size distribution of abalone populations, a pattern also present in the most recent survey in 2009(WDFW)(methods according to Rothaus et al. 2008)(Figure 3). This signifies a shift in abalone population age structure from younger to older animals, indicative of repeated recruitment failure (Rothaus et al. 2008). Recruitment failure following substantial declines in abalone density have been demonstrated elsewhere, for example in British Columbia, Canada (Tomascik and Holmes 2003) and in California (e.g., Miner et al. 2006). In Washington, the observed increases in mean shell length oppose the notion that the observed populations declines are a result of continued illegal harvest, because poaching is likely to result in a shift in length frequency toward smaller individuals (Rothaus et al. 2008). Pinto abalone populations may be unlikely to recover without intervention (Rothaus et al. 2008). Since 2004, a program of hatchery-based rearing and outplanting aimed at restoring abalone populations in Washington State has been led by the Puget Sound Restoration Fund (PSRF) and several local partners. In the summer of 2009, nearly 2,000 abalone were outplanted near Anacortes and Port Angeles, Washington.
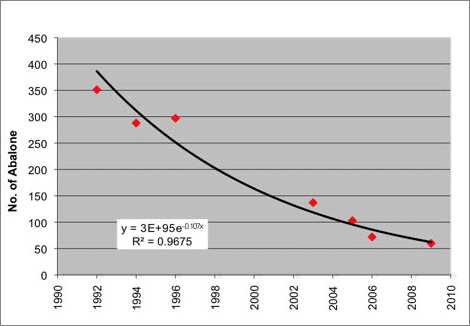
Figure 2. Pinto abalone abundance in the San Juan archipelago. Trends in abundance at 10 index stations from 1992 to 2009 (Figure produced by WDFW from unpublished data used with permission; methods according to Rothaus et al. 2008).
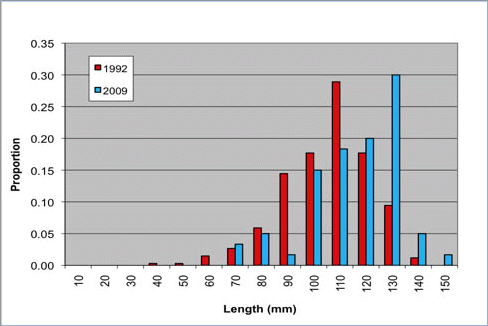
Figure 3. Pinto abalone shell length frequency in the San Juan archipelago. Trends in shell length from 10 index sites from 1992 to 2009 (Figure produced by WDFW from unpublished data used with permission; methods according to Rothaus et al. 2008).
Uncertainties
Many aspects of abalone biology and ecology are not well understood yet may be important in explaining both the decline and the recovery potential for pinto abalone in the Puget Sound region. While recreational fisheries likely played a role in the decline of pinto abalone in the San Juan Islands, the relative importance of harvesting and other factors is not known. While predation, habitat preferences, food availability and abiotic conditions will all likely affect the success of restoration efforts, the extent to which each of these factors may limit abalone populations is not well understood.
Summary
Pinto abalone are in severe decline in Puget Sound waters and are presently at densities where they may not be self-sustaining. Monitoring at index stations in the San Juan Islands showed an 83% decrease in abundance since 1992 despite their listing as federal species of concern, state candidate species, and the cessation of recreational harvest in 1994. Shell length surveys reveal that the population of pinto abalone in the San Juan Islands is aging without replacement although the direct causes of this recruitment failure warrant continued investigation. The long-term success of current hatchery-based rearing and outplanting programs is unknown at this time as efforts were recently initiated over the last five years.
Literature Cited
Babcock, R., and J. Keesing. 1999. Fertilization biology of the abalone Haliotis laevigata: laboratory and field studies. Canadian Journal of Fisheries and Aquatic Sciences 56:1668-1678.
Bargmann, G. G. 1984. Recreational diving in the State of Washington and the associated harvest of food fish and shellfish. State of Washington, Dept. of Fisheries, Marine Fish Program, Olympia, WA.
Bouma, J. V. 2007. Early life history dynamics of pinto abalone (Haliotis kamtschatkana) and implications for recovery in the San Juan archipelago, Washington State. Thesis (M S ). School of Aquatic and Fisheries Science, University of Washington, Seattle, WA.
Campell, A. E. 2000. Workshop on rebuilding abalone stocks in British Columbia. Pages 173-185 in Can. Spec. Publ. Fish. Aquat. Sci. .
COSEWIC. 2009. COSEWIC assessment and update status report on the Northern Abalone Haliotis kamtschatkana in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, CAN.
Miner, C. M., J. M. Altstatt, P. T. Raimondi, and T. E. Minchinton. 2006. Recruitment failure and shifts in community structure following mass mortality limit recovery prospects of black abalone. Marine Ecology-Progress Series 327:107-117.
Rothaus, D. P., B. Vadopalas, and C. S. Friedman. 2008. Precipitous declines in pinto abalone (Haliotis kamtschatkana kamtschatkana) abundance in the San Juan Archipelago, Washington, USA, despite statewide fishery closure. Canadian Journal of Fisheries and Aquatic Sciences 65:2703(2709).



