

Background
Rapid growth and accumulation of phytoplankton or other algae can cause algal blooms. Bloom-forming algae that have harmful effects on people or wildlife are commonly termed harmful algal blooms (HABs). In Puget Sound, HABs may be caused by phytoplankton such as dinoflagellates of the genus Alexandrium, diatoms of the genus Psuedo-nitzchia, raphidophytes of the genus Heterosigma or by ulvoid seaweeds. Suspension-feeding bivalves, such as mussels, clams and oysters, can accumulate biotoxins to dangerous levels during HAB events, leading to illness such as paralytic shellfish poisoning (PSP) or amnesic shellfish poisoning (ASP) when the shellfish are ingested by humans, marine mammals and marine birds (Nishitani and Chew 1984, Hallegraeff 1993).
The Washington Department of Health(WDOH) Office of Shellfish Safety and Water Protection regularly monitors biotoxin levels in both recreational and commercial shellfish areas in Puget Sound. The Washington State Public Health Laboratory supports the WDOH through the analysis of shellfish samples. When high levels are detected in sample tissues, shellfish harvest areas are closed in order to protect shellfish consumers from biotoxin-related illness. Closures can have significant effects on commercial, recreational, and subsistence harvest. Episodes of high biotoxin levels are currently unpredictable in time or space due to the interaction of multiple poorly understood environmental factors (Moore et al. 2009).
Paralytic Shellfish Poisoning
Seasonal restrictions on commercial and recreational shellfish harvest due to PSP, sometimes known as "red tide", are common in Washington. The biotoxin that causes PSP temporarily interferes with the transmission of nerve impulses in warm-blooded animals. Symptoms of PSP in humans range from nausea, vomiting, numbness of the lips and tongue and muscle paralysis to death by cardio-respiratory arrest. There is no known antidote for the toxin, and cooking does not destroy the toxin.
Several microscopic organisms that naturally exist in marine water produce the PSP toxin. The species that causes PSP in Washington marine waters is the dinoflagellate Alexandrium catenella (Determan et al. 2001). Alexandrium is typically present in small numbers; however, when environmental conditions are favorable, rapid reproduction and accumulation can occur, and shellfish can accumulate the toxin to dangerous levels during such bloom events (Zingone and Enevoldsen 2000, Moore et al. 2009).
WDOH closes areas for shellfish harvest when PSP toxin levels equal or exceed the Food and Drug Administration standard of 80 micrograms (μg) of toxin per 100 grams of shellfish tissue. Areas are not reopened until testing has confirmed that the PSP toxin has declined to a safe level. Butter clams may experience extended closures because they typically retain the PSP toxin longer than other shellfish (up to one year).
Sentinel Mussel Monitoring Program
The Sentinel Mussel Monitoring Program is an early warning system for marine biotoxins established by WDOH. Mussels generally register PSP toxin levels more quickly than other shellfish. Consequently, mussels are used as “sentinels” to determine whether PSP toxins are increasing in a given area. Under this monitoring program, mussels are placed in cages and set in strategic growing areas throughout Puget Sound. Mussel samples are then collected either biweekly or monthly and tested for levels of PSP. Rising PSP levels in these mussels trigger more targeted and frequent sampling regimens in other shellfish species in the affected area.
With assistance from local health jurisdictions, local tribes, the Puget Sound Restoration Fund, and volunteers, WDOH maintained and monitored 69 collection sites in 2008 (WDOH 2009). In addition to the sentinel mussel locations, commercial mussels were routinely monitored at Westcott Bay in San Juan Island and at Penn Cove in Whidbey Island.
Amnesic Shellfish Poisoning
Domoic acid is a naturally-occurring toxin produced by a species of microscopic marine diatoms of the genus Pseudo-nitzschia. The human illness known as ASP or domoic acid poisoning is caused by eating fish, shellfish or crab containing the toxin. ASP can result in gastrointestinal and neurological disorders within 24-48 hours of toxic shellfish consumption by humans, and can be life-threatening. There is no antidote for domoic acid poisoning and cooking does not destroy the toxin.
The razor clam and Dungeness crab fisheries on the outer coast of Washington State have incurred losses due to occurrences of domoic acid over the past two decades. In the fall of 1991, domoic acid was first detected in razor clams off the coast of Washington and caused several mild cases of ASP (Horner and Postel 1993). This prompted WDOH to begin monitoring all major shellfish growing areas for domoic acid. Research shows that razor clams accumulate domoic acid in the edible tissue (foot, siphon, and mantle) and are slow to rid themselves of the toxin (Wekell et al. 1994) due to the presence of a high affinity glutamate binding protein (Trainer and Bill 2004). However, razor clams can continue to function in marine environments with high concentrations of domoic acid (Trainer and Bill 2004), resulting in extended closures of shellfish beds of the outer coast of Washington. In Dungeness crab, domoic acid primarily accumulates in the viscera. The level of domoic acid determined to be unsafe for human consumption is 20 parts per million (ppm) in molluscan shellfish and 30 ppm for Dungeness crab viscera. Dungeness crab harvest areas are closed when three out of six individual crab viscera equals or exceeds 30 ppm.
Within Puget Sound, the first occurrence of domoic acid was in blue mussels harvested in Kilisut Harbor in 2003 (Bill et al. 2006), raising concerns about the possibility of shellfish closures similar to those on the outer coast. Puget Sound was presumed to be less susceptible to domoic acid closures due to the absence of harvested species (razor clams and Dungeness crab) that retain domoic acid for long periods. Many shellfish species that are harvested in Puget Sound, such as mussels, littleneck clams, and oysters, are able to depurate domoic acid over a period of hours or days (Novaczek 1992), whereas the ability of other species such as geoduck to retain or release domoic acid has not yet been determined (Trainer et al. 2007).
Heterosigma
While not responsible for illnesses in humans, blooms of the small, unicellular, flagellated raphidophyte Heterosigma akashiwo have been shown to kill fish through the likely production of neurotoxins that disrupt respiratory and osmoregulatory gill functions (Khan et al. 1997, Hard et al. 2000). Farmed fish are particularly susceptible to mortality from increased concentrations of Heterosigma (Chang et al. 1990, Hard et al. 2000). Increased water column stratification and high temperatures have both been correlated with Heterosigma blooms although the precise causes for blooms remain uncertain (Bearon et al. 2006, O'Halloran et al. 2006).
Ulvoids
Blooms of ulvoid seaweeds are manifested by large quantities of green algal biomass washing up on beaches where decomposition occurs or in seagrass beds where mortality of seagrass through smothering is possible (den Hartog 1994, Nelson and Lee 2001). The thin blade-like morphology of ulvoids is thought to contribute to their ability to respond quickly to favorable environmental conditions such as increased nutrients and light (e.g., Littler and Littler 1980). As such, they have can competitively displace other algal species and seagrasses (e.g., den Hartog 1994, Anderson et al. 1996, Valiela et al. 1997). While not typically associated with the production of toxins, there is emerging evidence that ulvoid algae can produce allelopathic compounds that are detrimental to the development and growth of invertebrate larvae and other algae (Nelson et al. 2003a, Van Alstyne et al. 2007). Two genera of ulvoid seaweeds are common in Puget Sound: Ulva (which includes the former genus Enteromorpha) and Ulvaria (formerly referred to as Monostroma)(Nelson et al. 2003b). Despite their morphological similarity, these genera differ ecologically. A combination of field and lab observations conducted by Nelson and colleagues has demonstrated that Ulva is more tolerant of desiccation stress, produces lower levels of allelopathic compounds and is found more commonly in intertidal habitats whereas Ulvaria is less tolerant of desiccation stress, produces higher levels of allelopathic compounds and is more commonly found in subtidal habitats (Nelson et al. 2003a, Nelson et al. 2003b, Nelson et al. 2008, Nelson et al. 2010).
Status
PSP and ASP
In 2008, only 12 of 2,798 samples (0.4%) of shellfish tested by the Washington State Public Health Laboratory detected levels of PSP toxins greater than 1,000 micrograms and no PSP-related illnesses in humans were reported (WDOH 2009) (Table 1). However, 23 subtidal geoduck clamtracts were closed due to elevated PSP toxin levels and two general closures for “all shellfish species” occurred. Notably, one geoduck tract closure included a recall of 3,368 lbs of geoduck clams. In 2008, the highest PSP levels in blue mussels were found in Mystery Bay, Kilisut Harbor (Table 1).
Table 1. Areas of highest PSP levels in 2008 (WDOH 2009)
Date | Harvest Area | Species | Toxin Level* |
08/10/2008 | Mystery Bay, Kilisut Harbor | Blue Mussel | 2,602 |
06/17/2008 | Semiahmoo Marina, Drayton Harbor | Blue Mussel | 1,831 |
08/11/2008 | Scow Bay, Kilisut Harbor | Blue Mussel | 1,779 |
09/25/2008 | Dockton, Quartermaster Harbor | Blue Mussel | 1,462 |
06/17/2008 | Birch Bay Village, Birch Bay | Blue Mussel | 1,456 |
08/06/2008 | Fort Flagler, Kilisut Harbor | Blue Mussel | 1,347 |
11/12/2008 | Ediz Hook, East Straits | Blue Mussel | 1,097 |
- micrograms per 100 grams of shellfish meat tissue
Approximately 12 Dungeness crab and 1,318 molluscan shellfish samples were tested by WDOH for domoic acid in 2008. The low sample size for crabs was driven by lack of toxin in the first 12 samples, which prompted a halt in further testing of Dungeness crab. There were no shellfish closures due to high levels of domoic acid in 2008, nor any reported ASP illnesses (WDOH 2009). The highest levels of domoic acid observed in Puget Sound molluscs in 2008 were at Squaxin Passage and Budd Inlet (Table 2).
Table 2. Areas of highest domoic acid levels in 2008 (WDOH 2009)
Date | Harvest Area | Species | Toxin Level* |
06/24/2008 | Squaxin Passage | Blue Mussel | 3 |
06/19/2008 | Budd Inlet | Blue Mussel | 3 |
01/07/2008 | Kalaloch Beach North | Razor Clam | 3 |
11/06/2008 | Long Beach Reserve | Razor Clam | 2 |
10/01/2008 | Sequim Bay | Blue Mussel | 2 |
06/24/2008 | South Tacoma Narrows | Blue Mussel | 2 |
- parts per million per 1 gram of shellfish meat tissue
Heterosigma
Heterosigma has been reported in various locations in Puget Sound and has been linked to fish mortality at fish farms in Puget Sound (Hershberger et al. 1997, Tyrrell et al. 2002). Despite the potential problem of financial damage to fish farms from Heterosigma or of mortality of wild fish from Heterosigma blooms, available data the spatial variation of both Heterosigma occurrence and the frequency of associated fish mortality events is limited.
Ulvoids
A study conducted by Nelson et al. (2003b) assessing biomass of ulvoids in locations in Puget Sound (Figure 1) in summer of 2000 found that the species composition, depth, and abundance of ulvoids was variable throughout Puget Sound (Figure 2). In a more detailed analysis linking ulvoid biomass to abiotic variables on the coast of Blakely Island in the San Juan Archipelago, Nelson et al. (2003b) found that increased biomass was positively correlated with increased nitrogen, a finding that is consistent with studies conducted in other locations (e.g., Sfriso et al. 1992, Anderson et al. 1996).

Figure 1. Sampling locations for ulvoid algae conducted by Nelson et al.(2003b)(Reprinted with permission from Botanica Marina and De Gruyter Publishing).
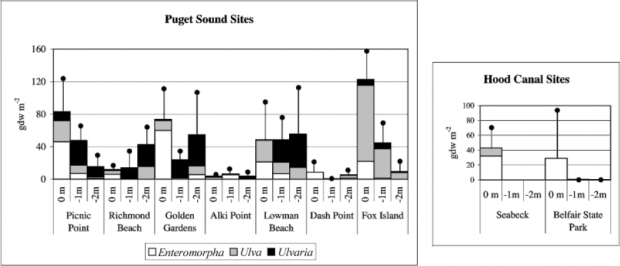
Figure 2. Biomass (mean + 1SD) by genus and by depth if ulvoid algae at locations throughout Puget Sound (Nelson et al. 2003b)(Reprinted with permission from Botanica Marina and De Gruyter Publishing).
Trends
PSP
Harmful algal blooms of Alexandrium were widespread and prevalent in the northern regions of Puget Sound (e.g., Sequim and Discovery Bays) in the 1950s and 1960s, but extended southward in the 1970s and 1980s to inner regions of Puget Sound (Trainer et al. 2003). More recent occurrences of PSP toxins in Washington shellfish and crab have been variable. Although high levels of PSP were detected in many years between 1990 and 2006, in some years (e.g., 1995, 2007, 2008) PSP toxin levels remained low (Figure 3). Despite this variability, the frequency of instances of high levels of PSP toxins detected by WDOH monitoring in Washington State has increased since 1957(Figure 3), a trend that is consistent with a worldwide increase in PSP toxic events since the 1950s (Nishitani and Chew 1984, Hallegraeff 1993, Trainer et al. 2003, Maso and Garces 2006).
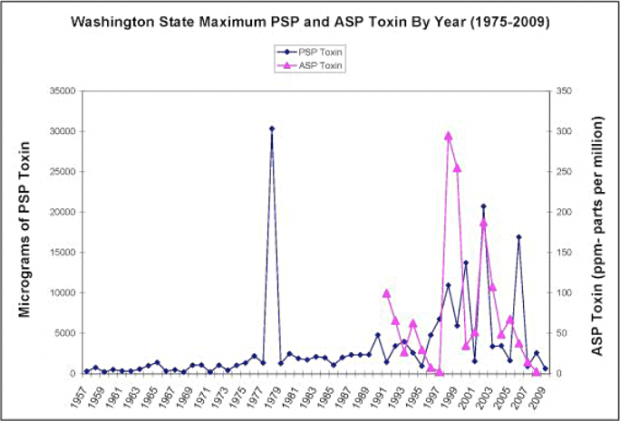
Figure 3. Annual maximum concentrations of PSP and ASP toxins observed in Washington State (WDOH, reprinted from Puget Sound Partnership 2009).
Moore et al. (2009) caution the use of PSP levels in shellfish tissues as a proxy for algal cell density in the water column due to the difference in accumulation and depuration rates of shellfish species. To investigate trends and possible relationships with large-scale climate and local environmental factors, Moore et al. (2009) analyzed PSP levels in blue mussels between 1993 and 2007. A combination of warm air and water temperatures and low streamflow appears to be favorable for PSP toxin accumulation in mussels, but advanced warning of events may be constrained by the same factors as for weather prediction, and is therefore limited to approximately one to two weeks (Moore et al. 2008, Moore et al. 2009). No increase in the frequency, magnitude, duration, or geographic scope of HAB events was detected, yet a significant basin-wide trend for closures to be imposed earlier in the year was observed over the period.
ASP
Blooms of Pseudo-nitzchia continue to affect Washington’s outer coast since the first fisheries closure due to ASP toxins in 1991. Exceptional years of domoic acid-associated beach, razor clam, and Dungeness crab closures in Washington include 1991, 1998-1999, 2002-2003, and 2005 (Horner and Postel 1993, Trainer et al. 2007)(Figure 3). The prolonged closures of 1998-1999 and 2002-2003 (>1.5 years) resulted in significant commercial, recreational, and tribal shellfish harvest losses in Washington State (Dyson and Huppert in press, corrected proof). Out of concern for ASP toxins in the highly populated Puget Sound region, WDOH has monitored throughout Puget Sound since 1991. Pseudo-nitzschia blooms were reported in Puget Sound in 2003 and 2005, causing concern that blooms could impact the valuable fisheries there (Trainer et al. 2009).
Heterosigma
The bloom-forming alga Heterosigma akashiwo is recognized as a potential problem in Puget Sound. Despite a number of current studies on this HAB-forming alga, data are not yet available to determine spatial and temporal trends in Heterosigma abundances or the frequency of toxic events in Puget Sound.
Ulvoids
Published accounts of temporal trends in ulvoid abundances in Puget Sound are lacking. At least one investigation currently is underway to estimate ulvoid abundance from archival video surveys.
Uncertainties
Trend analysis of harmful algal blooms is difficult due to the lack of understanding about the dynamics that drive them, although this is an area of active research (e.g., Bearon et al. 2006, Nelson et al. 2008, Moore et al. 2009). Environmental conditions such as circulation, temperature, sunlight, nutrients, and salinity as well as the presence of algal predators, parasites and algal disease organisms all likely play a role in the formation, magnitude, and persistence of blooms. While PSP and ASP toxin levels currently are monitored and reported by WDOH, published data regarding spatial and temporal trends in Heterosigma and ulvoid abundances in Puget Sound are lacking.
Summary
Harmful algal blooms in Puget Sound have been variable over the past two decades, but appear to be increasing since WDOH began monitoring in 1957. Current monitoring efforts are not sufficient to provide accurate forecasting of ASP and PSP-related bloom events beyond one to two weeks, but forecasting could be improved by increased temporal and spatial scale and automated devices. While there is emerging concern about blooms of Heterosigma and ulvoids, , data that address these concerns currently is lacking for Puget Sound.
References
Anderson, R. J., P. M. S. Monteiro, and G. J. Levitt. 1996. The effect of localised eutrophication on competition between Ulva lactuca (Ulvaceae, Chlorophyta) and a commercial resource of Gracilaria verrucosa (Gracilariaceae, Rhodophyta). Hydrobiologia 327:291-296.
Bearon, R. N., D. Grunbaum, and R. A. Cattolico. 2006. Effects of salinity structure on swimming behavior and harmful algal bloom formation in Heterosigma akashiwo, a toxic raphidophyte. Marine Ecology-Progress Series 306:153-163.
Bill, B. D., F. H. Cox, R. A. Horner, J. A. Borchert, and V. L. Trainer. 2006. The first closure of shellfish harvesting due to domoic acid in Puget Sound, Washington, USA. African Journal of Marine Science 28:435-440.
Chang, F. H., C. Anderson, and N. C. Boustead. 1990. 1st Record of a Heterosigma (Raphidophyceae) bloom with associated mortality of cage-reared salmon in Big Glory Bay, New Zealand New Zealand Journal of Marine and Freshwater Research 24:461-469.
den Hartog, C. 1994. Suffocation of a littoral Zostera bed by Enteromorpha radiata. Aquatic Botany 47:21-28.
Determan, T. A., S. Washington . Office of Food, P. Shellfish, and P. Puget Sound Ambient Monitoring. 2001. Paralytic shellfish poisoning (PSP) patterns in Puget Sound shellfish, year 2000 : a report for the Puget Sound Ambient Monitoring Program. Office of Food Safety and Shellfish Programs, Washington State Dept. of Health, [Olympia, Wash.].
Dyson, K., and D. D. Huppert. in press, corrected proof. Regional economic impacts of razor clam beach closures due to harmful algal blooms (HABs) on the Pacific coast of Washington. Harmful Algae In Press, Corrected Proof.
Hallegraeff, G. 1993. A review of harmful algae blooms and their apparent global increase. Phycologia 32:79 - 99.
Hard, J. J., L. Connell, W. K. Hershberger, and L. W. Harrell. 2000. Genetic variation in mortality of chinook salmon during a bloom of the marine alga Heterosigma akaskiwo. Journal of Fish Biology 56:1387-1397.
Hershberger, P. K., J. E. Rensel, A. L. Matter, and F. B. Taub. 1997. Vertical distribution of the chloromonad flagellate Heterosigma carterae in columns: implications for bloom development. Canadian Journal of Fisheries and Aquatic Sciences 54:2228-2234.
Horner, R. A., and J. R. Postel. 1993. Toxic diatoms in western Washington waters (US West coast). Hydrobiologia 269:197-205.
Khan, S., O. Arakawa, and Y. Onoue. 1997. Neurotoxins in a toxic red tide of Heterosigma akashiwo (Raphidophyceae) in Kagoshima Bay, Japan. Aquaculture Research 28:9-14.
Littler, M. M., and D. S. Littler. 1980. The evolution of thallus form and survival strategies in benthic marine macroalgae: Field and laboratory tests of a functional form model. The American Naturalist 116:25.
Maso, M., and E. Garces. 2006. Harmful microalgae blooms (HAB); problematic and conditions that induce them. Marine Pollution Bulletin 53:620-630.
Moore, S., V. Trainer, N. Mantua, M. Parker, E. Laws, L. Backer, and L. Fleming. 2008. Impacts of climate variability and future climate change on harmful algal blooms and human health. Environmental Health 7:S4.
Moore, S. K., N. J. Mantua, B. M. Hickey, and V. L. Trainer. 2009. Recent trends in paralytic shellfish toxins in Puget Sound, relationships to climate, and capacity for prediction of toxic events. Harmful Algae 8:463-477.
Nelson, T. A., K. Haberlin, A. V. Nelson, H. Ribarich, R. Hotchkiss, K. L. V. Alstyne, L. Buckingham, D. J. Simunds, and K. Fredrickson. 2008. Ecological and physiological controls of species composition in green macroalgal blooms. Ecology 89:1287-1298.
Nelson, T. A., and A. Lee. 2001. A manipulative experiment demonstrates that blooms of the macroalga Ulvaria obscura can reduce eelgrass shoot density. Aquatic Botany 71:149-154.
Nelson, T. A., D. J. Lee, and B. C. Smith. 2003a. Are "green tides" harmful algal blooms? Toxic properties of water-soluble extracts from two bloom-forming macroalgae, Ulva fenestrata and Ulvaria obscura (Ulvophyceae). Journal of Phycology 39:874-879.
Nelson, T. A., A. V. Nelson, and M. Tjoelker. 2003b. Seasonal and spatial patterns of "Green tides" (Ulvoid algal blooms) and related water quality parameters in the coastal waters of Washington state, USA. Botanica Marina 46:263-275. De Gruyter (Publisher).
Nelson, T. A., J. Olson, L. Imhoff, and A. V. Nelson. 2010. Aerial exposure and desiccation tolerances are correlated to species composition in "green tides'' of the Salish Sea (northeastern Pacific). Botanica Marina 53:103-111.
Nishitani, L., and K. K. Chew. 1984. Recent developments in paralytic shellfish poisoning research. Aquaculture 39:317-329.
Novaczek, I., Madhyastha, M.S., Ablett, R.F., Donald, A., Johnson, G., Nijjar, M.S., Sims, D.E. 1992. Depuration of domoic acid from live blue mussels (Mytilus edulis). Can. J. Fish. Aquat. Sci. 49:312-318.
O'Halloran, C., M. W. Silver, T. R. Holman, and C. A. Scholin. 2006. Heterosigma akashiwo in central California waters. Harmful Algae 5:124-132.
Puget Sound Partnership. 2009. Ecosystem Status & Trends. Puget Sound Partnership, Seattle, WA.
Sfriso, A., B. Pavoni, A. Marcomini, and A. Orio. 1992. Macroalgae, nutrient cycles, and pollutants in the Lagoon of Venice. Estuaries and Coasts 15:517-528.
Trainer, V., B.-T. Eberhart, J. Wekell, N. Adams, L. Hanson, F. Cox, and J. Dowell. 2003. Paralytic shellfish toxins in Puget Sound, Washington State. J Shellfish Res 22:213 - 223.
Trainer, V. L., and B. D. Bill. 2004. Characterization of a domoic acid binding site from Pacific razor clam. Aquatic Toxicology 69:125-132.
Trainer, V. L., W. P. Cochlan, A. Erickson, B. D. Bill, F. H. Cox, J. A. Borchert, and K. A. Lefebvre. 2007. Recent domoic acid closures of shellfish harvest areas in Washington State inland waterways. Harmful Algae 6:449-459.
Tyrrell, J. V., L. B. Connell, and C. A. Scholin. 2002. Monitoring for Heterosigma akashiwo using a sandwich hybridization assay. Harmful Algae 1:205-214.
Valiela, I., J. McClelland, J. Hauxwell, P. J. Behr, D. Hersh, and K. Foreman. 1997. Macroalgal blooms in shallow estuaries: Controls and ecophysiological and ecosystem cnsequences. Limnology and Oceanography 42:1105-1118.
Van Alstyne, K., L. Koellermeier, and T. Nelson. 2007. Spatial variation in dimethylsulfoniopropionate (DMSP) production in Ulva lactuca (Chlorophyta) from the Northeast Pacific. Marine Biology 150:1127-1135.
WDOH, W. D. o. H. 2009. 2008 Annual Report: Commercial and Recreational Shellfish Areas in Washington State. Office of Shellfish and Water Protection.
Wekell, J. C., J. R. Gauglitz, E.J.Barnett, H.J.Hatfield, C.L.Simons, and D. Ayres. 1994. Occurrence of domoic acid in Washington state razor clams (Siliqua patula) during 1991–1993. Natural Toxins 2:197-205.
Zingone, A., and H. O. Enevoldsen. 2000. The diversity of harmful algal blooms: a challenge for science and management. Ocean & Coastal Management 43:725-748.


