

Chinook salmon and estuary use in Puget Sound

For more information, view the original reports: The role of estuaries in the ecology of adult Pacific salmon and trout in Puget Sound and The role of estuaries in the ecology of juvenile Pacific salmon and trout in Puget Sound.
Overview
Chinook salmon are the largest Pacific salmon as adults, and are highly valued for their symbolism, in commercial and recreational fisheries, and as a critical food source for endangered southern resident killer whales. Regrettably, the populations of wild Chinook salmon are in jeopardy, and the Puget Sound population complex is listed as Threatened under the ESA. Consequently, they are the subject of considerable research, monitoring, and restoration efforts.
Adults
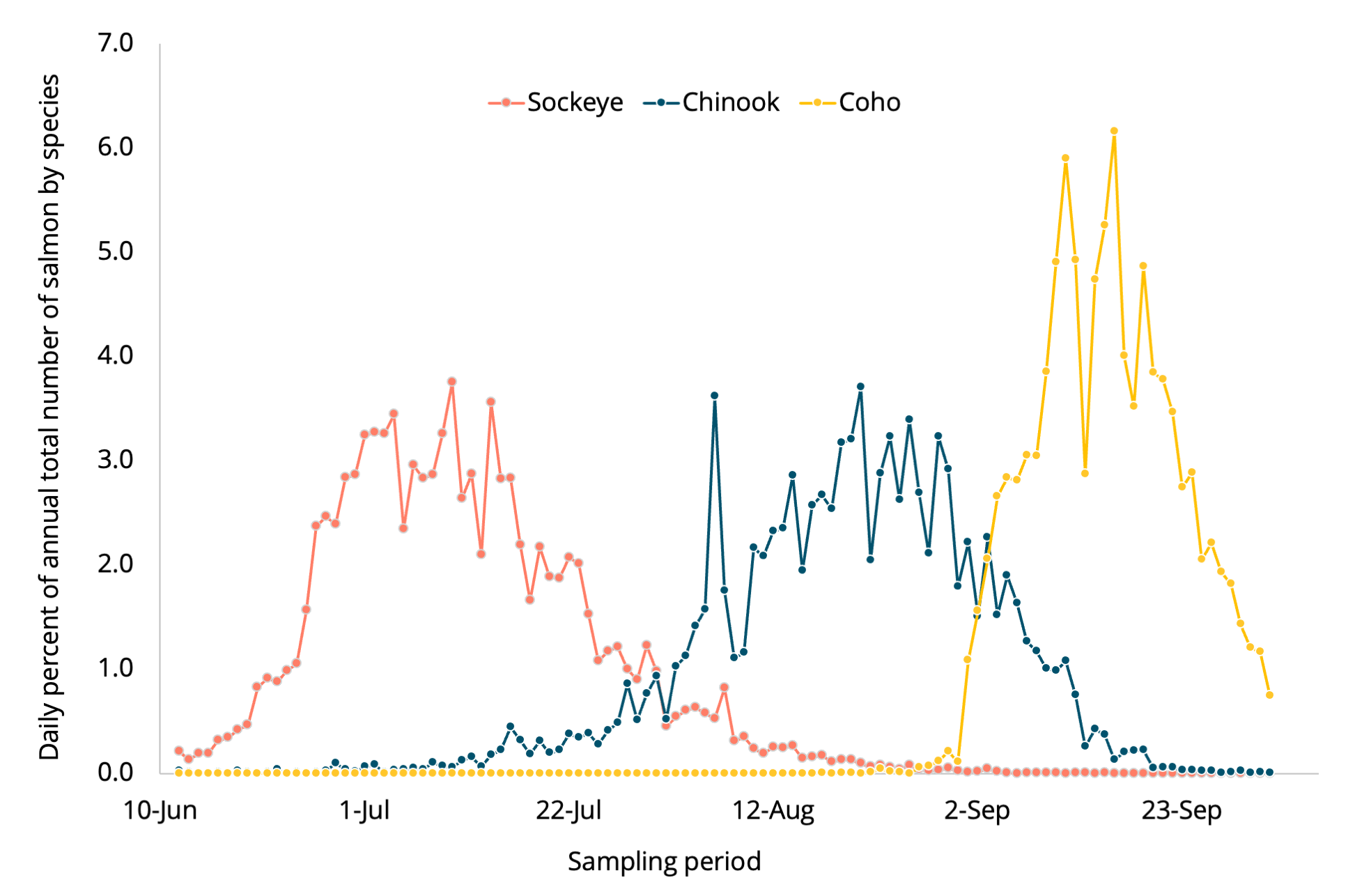
Unlike the relatively narrow migration timing of sockeye salmon, Chinook salmon pass through estuaries and up rivers over a much wider period, from spring to fall. Adults ascend the Sacramento River in every month of the year and in the Columbia River their counts at Bonneville Dam extend from early March to early December. Their runs are designated by the season when they leave marine waters (e.g., spring, summer and fall) and all these spawn in the fall, though the Sacramento River winter-run Chinook salmon spawn in the late spring. In Puget Sound, the great majority of Chinook salmon are fall-run, leaving marine waters in the late summer and early fall as water temperatures are peaking and dropping, and spawning in the fall.
Hatchery operations have tended to advance the timing of spawning, for example in the Issaquah Creek and University of Washington populations in the Lake Washington Basin, the Soos Creek population in the Green River (Quinn et al. 2002), and the Skagit River (Austin et al. 2021). In addition to the effects of hatchery programs on timing, there have also been some extirpations, so the current diversity in timing does not fully represent historical patterns, but spring, summer, and fall runs persist in Puget Sound, including spring runs in the Skagit and Puyallup rivers (Myers et al. 1998, Ruckelshaus et al. 2006).
How we assess the use of Puget Sound’s estuaries by Chinook salmon depends on how we view Puget Sound and the Salish Sea as a whole. In many ways Puget Sound is an estuary, and it is critical for Chinook salmon. Scientists knew that Chinook and coho salmon were routinely taken throughout the year in Puget Sound and doubtless the indigenous peoples knew this long before (Jordan and Evermann 1896). Some smolts leave and feed along the Pacific Ocean coast whereas others remain in Puget Sound until they mature, and there are even a few that go back and forth between the ocean and Puget Sound (Kagley et al. 2017).
The tendency to remain as “residents” in Puget Sound is not well understood but it occurs in many populations rather than being unique to one or a few (Chamberlin et al. 2011). Moreover, the residents do not move widely throughout Puget Sound but seem to spend most of their time in a limited area, based on data from 1976–2017, while others in their population migrate up the coast to British Columbia and even Alaska (Quinn et al. 2005). These residents, known as “blackmouth” by anglers in reference to the black gum line, remain important in recreational fisheries (Photo 1), including the Tengu Derby, held for many decades in the winter in Elliott Bay near downtown Seattle (Quinn et al. 2022). Thus, some Chinook salmon feed within Puget Sound, including waters near the mouths of rivers such as Elliott Bay and the Duwamish River, for their entire marine residence period. Estimates vary but the fraction of Puget Sound origin Chinook showing this resident pattern is large (O’Neill and West 2009), and perhaps they even make up the majority, though the proportion seems to vary among years (Chamberlin et al. 2011, Quinn and Losee 2022).

Resident Chinook feed actively while in Puget Sound, primarily on invertebrates when they enter marine waters and occupy the nearshore area, shifting to other invertebrates and fishes as they grow and move offshore (Duffy et al. 2010).Their diet is more dominated by fishes compared to the other Pacific salmon species (Beacham 1986, Quinn 2018), and this tendency increases as they grow (Prakash 1962, Beacham 1986, Daly et al. 2009). Consistent with this overall tendency, sub-adult Chinook salmon in Puget Sound prey heavily on fishes, and especially Pacific herring (Kirkness 1948, Healey 1976, Chamberlin et al. 2021), thus healthy and diverse herring populations are important for Chinook salmon. In addition to these residents, many Chinook salmon feed along the coast, primarily off British Columbia but as far north as Alaska and also off Oregon. These salmon pass through the Salish Sea on their homeward journey, and they travel in the upper 30 m of the water column, migrating about 1–2 km/hour (Hendriks 2024).
As they approach sexual maturity Chinook salmon ascend their natal rivers, but their progress up free-flowing rivers in our region has not been studied in detail. Those ascending the locks on their way into the Lake Washington basin (Figure 1) have an unusual set of challenges because the temperature, salinity, and dissolved oxygen change abruptly from the marine to freshwater environments. Chinook salmon have been studied in these habitats (Goetz and Quinn 2019), but it is not clear how their behavior informs us about those in the more natural estuaries. We might expect the early running spring Chinook to move upriver quickly to reach cool waters near their spawning sites where they can hold while completing sexual maturation. In contrast, the fall Chinook enter in a more advanced state of maturity and might spend more time in estuaries. However, this is really a hypothesis in need of testing rather than a general rule. To my knowledge, there is no specific, thorough study of estuarine behavior of different Chinook salmon runs in the same river.
In addition to the broad seasonal patterns of Chinook salmon timing, there is also the question of what factors cue them to migrate upriver. There is a common impression that increasing flows stimulate their migration but there is evidence to the contrary from the Klamath River (Strange 2010). In some cases, diversions of water for human uses result in unnaturally steady flows, and efforts to stimulate upstream migrating Chinook salmon with experimental pulsed flows has been either not efficacious (Hasler et al. 2014) or only slightly successful (Peterson et al. 2017). Overall, the factors controlling the upriver migrations of salmon have long been studied but remain poorly understood. Temperature, flow, and physical factors play a large role in some cases (rivers and salmon species) and a much smaller one in others, and the social interactions among salmon likely play a role as well (Berdahl et al. 2017).
Chinook salmon tend to migrate up large rivers and there is seldom, if ever, too little water in these rivers for them to ascend, and so how long they stay in estuaries may depend on other factors related to their internal condition (maturation, feeding, salt regulation, etc.) and the various environmental conditions there such as salinity, flow, temperature, and dissolved oxygen (Fujioka 1970). In particular, combinations of high temperature and low dissolved oxygen levels, as can occur in late summer, may stall upriver migration (Alabaster 1988, 1989). Moreover, the presence of predatory marine mammals may also affect how long Chinook and other salmon species stay in estuaries (Strange 2013).
Juveniles
Chinook juvenile life history and use of estuaries is complicated, because they migrate to salt water over larger ranges of sizes and dates than the other species. Juvenile Chinook salmon are classified as ocean-type if they migrate to the ocean in their first year of life, and stream-type if they feed for a whole year in the stream and migrate in their second spring of life (Gilbert 1913). The Puget Sound populations are dominated by ocean-type fish, and the stream-type is most common in higher elevation streams fed primarily by snowmelt such as the upper Sauk River system (Beechie et al. 2006).
The juvenile stage has an important connection to downstream migration timing, body size, and use of estuaries. The stream-type fish migrate in the spring, with generally similar timing to coho salmon after a year of feeding in rivers. They are large (ca. 100 – 140 mm),migrate directly downstream, and seem to move quickly from Puget Sound (Rice et al. 2011). In contrast, ocean-type juveniles may migrate downstream more or less immediately after they emerge from gravel nests at about 36-40 mm in length (“fry migrants”), or after feeding for several months in the river (“parr migrants”) at about 60 – 90 mm in length (Anderson and Topping 2018). Consequently, juvenile Chinook salmon enter estuaries over several months, and over a range of sizes (Chamberlin et al. 2022).

Superficially, it is not clear if a 60 mm fish caught in an estuary just arrived or if it arrived weeks earlier and at a smaller size and then grew. Chemical analysis of stable isotopes, however, can be used to infer the duration of residency by individual fish. Moore et al. (2016) conducted such analysis for salmon caught in the estuary of the Skeena River, B. C. and estimated that 25% of the juvenile Chinook salmon had spent at least 33 days there. Coho and pink salmon also seemed to use the estuary for feeding whereas sockeye salmon moved through quickly.
Because of their range of sizes and dates, juvenile Chinook salmon make use of a wide variety of estuarine habitats, and perhaps the tidal portions of rivers are especially important for them as they begin the transition to seawater tolerance. The study of Chinook salmon in estuaries is complicated not only by the range of size and dates when they enter estuaries but by the high proportion of hatchery-origin fish in many rivers. These fish are typically larger and less variable in size than wild fish, and also less variable in timing as they are typically released from the hatchery over a narrow window in time. As a consequence of these differences, and perhaps others as well, wild and hatchery Chinook salmon may differ in patterns of habitat use, as seen in the Skagit River estuary (Beamer et al. 2024), and also in diet, as shown in the Nisqually River estuary (Davis et al. 2018). In general, the hatchery-origin fish move though the estuary more rapidly than do the wild ones.
After entering the Salish Sea, many Chinook smolts migrate to the ocean coast of British Columbia, Washington, and to a lesser extent Oregon and Alaska (Labelle et al. 1997, Quinn et al. 2005, Beamish et al. 2008, Weitkamp 2012). However, it has also been known for more than a century that some Chinook salmon are present in the Salish Sea throughout the year. Based on analysis of coded wire tagging (CWT) data, Puget Sound is largely occupied by Chinook salmon (Weitkamp 2010) that originated there rather than elsewhere in the Salish Sea or more distant locations.
Before these fish get big enough to be caught in fisheries, they seem to move around within Puget Sound. For example, sampling in the Nisqually River estuary in August and September revealed juvenile Chinook salmon from three marine basins, nine Puget Sound rivers, and 14 hatcheries (Hayes et al. 2019), though the majority were from the Nisqually River and nearby populations. Such movement to non-natal estuaries is important to understand, as it means that things done to restore or degrade one estuary affect other populations as well. Similarly, studies revealed that 32 of 63 small streams draining into the Whidbey Basin of northern Puget Sound had juvenile Chinook salmon in the spring, but none of the streams supported spawning (Beamer et al. 2013).
In addition to the broad seasonal patterns of Chinook salmon timing, there is also the question of what factors cue them to migrate upriver. There is a common impression that increasing flows stimulate their migration but there is evidence to the contrary from the Klamath River (Strange 2010). In some cases, diversions of water for human uses result in unnaturally steady flows, and efforts to stimulate upstream migrating Chinook salmon with experimental pulsed flows has been either not efficacious (Hasler et al. 2014) or only slightly successful (Peterson et al. 2017). Overall, the factors controlling the upriver migrations of salmon have long been studied but remain poorly understood. Temperature, flow, and physical factors play a large role in some cases (rivers and salmon species) and a much smaller one in others, and the social interactions among salmon likely play a role as well (Berdahl et al. 2017).
Chinook salmon tend to migrate up large rivers and there is seldom, if ever, too little water in these rivers for them to ascend, and so how long they stay in estuaries may depend on other factors related to their internal condition (maturation, feeding, salt regulation, etc.) and the various environmental conditions there such as salinity, flow, temperature, and dissolved oxygen (Fujioka 1970). In particular, combinations of high temperature and low dissolved oxygen levels, as can occur in late summer, may stall upriver migration (Alabaster 1988, 1989). Moreover, the presence of predatory marine mammals may also affect how long Chinook and other salmon species stay in estuaries (Strange 2013).
References
Alabaster, J. S. 1988. The dissolved oxygen requirements of upstream migrant chinook salmon, Oncorynchus tshawytscha, in the lower Willamette River, Oregon. Journal of Fish Biology 32:635-636.
Alabaster, J. S. 1989. The dissolved oxygen and temperature requirements of king salmon, Oncorhynchus tshawytscha, in the San Joaquin Delta, California. Journal of Fish Biology 34:331-332.
Anderson, J. H. and P. C. Topping. 2018. Juvenile life history diversity and freshwater productivity of Chinook Salmon in the Green River, Washington. North American Journal of Fisheries Management 38:180-193.
Austin, C. S., T. E. Essington, and T. P. Quinn. 2021. In a warming river, wild Chinook salmon spawn later but hatchery-origin conspecifics do not. Canadian Journal of Fisheries and Aquatic Sciences 78:68-77.
Beacham, T. D. 1986. Type, quantity, and size of food in Pacific salmon (Oncorhynchus) in the Strait of Juan de Fuca, British Columbia. Fishery Bulletin 84:77-89.
Beamer, E. M., C. M. Greene, J. Chamberlin, W. G. Hood, C. Ellings, S. Hodgson, and T. Zackey. 2024. Landscape determinants of aquatic estuarine habitat use by juvenile Chinook salmon. Canadian Journal of Fisheries and Aquatic Sciences 81:747-767.
Beamer, E. M., W. T. Zackey, D. Marks, D. Teel, D. Kuligowski, and R. Henderson. 2013. Juvenile Chinook salmon rearing in small non-natal streams draining into the Whidbey Basin. LaConner, WA.
Beamish, R. J., R. M. Sweeting, K. L. Lange, and C. M. Neville. 2008. Changes in the population ecology of hatchery and wild coho salmon in the Strait of Georgia. Transactions of the American Fisheries Society 137:503-520.
Beechie, T., E. Buhle, M. Ruckleshaus, A. Fullerton, and L. Holsinger. 2006. Hydrologic regime and the conservation of salmon life history diversity. Biological Conservation 130:560-572.
Berdahl, A., P. A. H. Westley, and T. P. Quinn. 2017. Social interactions shape the timing of spawning migrations in an anadromous fish. Animal Behaviour 126:221-229.
Chamberlin, J., E. Petrou, W. Duguid, R. Barsh, F. Juanes, J. Qualley, and L. Hauser. 2021. Phenological diversity of a prey species supports life-stage specific foraging opportunity for a mobile consumer. ICES Journal of Marine Science 78:3089-3100.
Chamberlin, J. W., J. Hall, W. T. Zackey, F. Leonetti, M. Rustay, and C. Rice. 2022. Spatial and temporal variability in Chinook salmon abundance reflects opportunity to support life history diversity in an estuarine landscape. Estuaries and Coasts 45:882-896.
Chamberlin, J. W., T. E. Essington, J. W. Ferguson, and T. P. Quinn. 2011. The influence of hatchery rearing practices on salmon migratory behavior: Is the tendency of Chinook salmon to remain within Puget Sound affected by size and date of release? . Transactions of the American Fisheries Society 140:1398-1408.
Daly, E. A., R. D. Brodeur, and L. A. Weitkamp. 2009. Ontogenetic shifts in diets of juvenile and subadult coho and Chinook salmon in coastal marine waters: important for marine survival? Transactions of the American Fisheries Society 138:1420-1438.
Davis, M. J., I. Woo, C. S. Ellings, S. Hodgson, D. A. Beauchamp, G. Nakai, and S. E. W. De La Cruz. 2018. Integrated diet analyses reveal contrasting trophic niches for wild and hatchery juvenile Chinook Salmon in a large river delta. Transactions of the American Fisheries Society 147:818--841.
Duffy, E. J., D. A. Beauchamp, R. M. Sweeting, R. J. Beamish, and J. S. Brennan. 2010. Ontogenetic diet shifts of juvenile Chinook salmon in nearshore and offshore habitats of Puget Sound. Transactions of the American Fisheries Society 139:803-823.
Fujioka, J. T. 1970. Possible effects of low dissolved oxygen content in the Duwamish River estuary on migrating adult chinook salmon. University of Washington, Seattle, WA.
Gilbert, C. H. 1913. Age at maturity of the Pacific coast salmon of the genus Oncorhynchus. Bulletin of the Bureau of Fisheries 32:1-22.
Goetz, F. A. and T. P. Quinn. 2019. Behavioral thermoregulation by adult Chinook salmon (Oncorhynchus tshawytscha) in estuary and freshwater habitats prior to spawning. Fishery Bulletin 117:258-271.
Hasler, C. T., E. Guimond, B. Mossop, S. G. Hinch, and S. J. Cooke. 2014. Effectiveness of pulse flows in a regulated river for inducing upstream movement of an imperiled stock of Chinook salmon. Aquatic Sciences 76:231--241.
Hayes, M. C., S. Hodgson, C. S. Ellings, W. D. Duval, and S. P. Rubin. 2019. Seasonal use of a nonnatal marine basin by juvenile hatchery Chinook Salmon. Marine and Coastal Fisheries 11:437-453.
Healey, M. C. 1976. Herring in the diets of Pacific salmon in Georgia Strait. Fisheries Research Board of Canada Manuscript Report 1382:1-38.
Hendriks, B. J. L. 2024. Behaviour and movement of return migrating adult Chinook salmon (Oncorhynchus tshawytscha) through the Salish Sea. MSc thesis. University of British Columbia, Vancouver, B. C.
Jordan, D. S. and B. W. Evermann. 1896. The Fishes of North and Middle America. Smithsonian Institution, Washington, D. C.
Kagley, A. N., J. M. Smith, K. L. Fresh, K. E. Frick, and T. P. Quinn. 2017. Residency, partial migration, and late egress of sub-adult Chinook salmon (Oncorhynchus tshawytscha) and comparisons with coho salmon (O. kisutch) in Puget Sound, Washington. Fishery Bulletin 115:544-555.
Kirkness, W. 1948. Food of the chinook and silver salmon of Puget Sound. Annual Report, Washington Department of Fisheries 1948:28-31.
Labelle, M., C. J. Walters, and B. Riddell. 1997. Ocean survival and exploitation of coho salmon (Oncorhynchus kisutch) stocks from the east coast of Vancouver Island, British Columbia. Canadian Journal of Fisheries and Aquatic Sciences 54:1433-1449.
Moore, J. W., J. Gordon, C. Carr-Harris, A. S. Gottesfeld, S. M. Wilson, and J. H. Russell. 2016. Assessing estuaries as stopover habitats for juvenile Pacific salmon. Marine Ecology Progress Series 559:201-215.
Myers, J. M., R. G. Kope, G. J. Bryant, D. Teel, L. J. Lierheimer, T. C. Wainwright, W. S. Grant, F. W. Waknitz, K. Neely, S. T. Lindley, and R. S. Waples. 1998. Status review of chinook salmon from Washington, Idaho, Oregon and California. NOAA Technical Memorandum NMFS-NWFSC-35, National Oceanic and Atmospheric Administration, National Marine Fisheries Service.
O'Neill, S. M. and J. E. West. 2009. Marine distribution, life history traits, and the accumulation of polychlorinated biphenyls in Chinook salmon from Puget Sound, Washington. Transactions of the American Fisheries Society 138:616-632.
Peterson, M. L., A. N. Fuller, and D. Demko. 2017. Environmental factors associated with the upstream migration of fall-run Chinook Salmon in a regulated river. North American Journal of Fisheries Management 37:78-93.
Prakash, A. 1962. Seasonal changes in feeding of coho and chinook (spring) salmon in southern British Columbia waters. Journal of the Fisheries Research Board of Canada 19:851-866.
Quinn, T. P. 2018. The Behavior and Ecology of Pacific Salmon and Trout, second edition. University of Washington Press, Seattle.
Quinn, T. P., B. R. Dickerson, and L. A. Vøllestad. 2005. Marine survival and distribution patterns of two Puget Sound hatchery populations of coho (Oncorhynchus kisutch) and chinook (O. tshawytscha) salmon. Fisheries Research 76:209-220.
Quinn, T. P. and J. P. Losee. 2022. Diverse and changing use of the Salish Sea by Pacific salmon, trout, and char. Canadian Journal of Fisheries and Aquatic Sciences 79:1003-1021.
Quinn, T. P., P. McGinnity, and T. E. Reed. 2016. The paradox of 'premature migration' by adult anadromous salmonid fishes: Patterns and hypotheses. Canadian Journal of Fisheries and Aquatic Sciences 73:1015-1030.
Quinn, T. P., J. A. Peterson, V. Gallucci, W. K. Hershberger, and E. L. Brannon. 2002. Artificial selection and environmental change: countervailing factors affecting the timing of spawning by coho and chinook salmon. Transactions of the American Fisheries Society 131:591-598.
Quinn, T. P., M. D. Scheuerell, J. P. Losee, and D. Hanada. 2022. Multi-decadal trends in body size of Puget Sound Chinook Salmon: Analysis of data from the Tengu Derby, a culturally unique fishery. Marine and Coastal Fisheries.
Rice, C. A., C. M. Greene, P. Moran, D. J. Teel, D. R. Kugliowski, R. R. Reisenbichler, E. M. Beamer, J. R. Karr, and K. L. Fresh. 2011. Abundance, stock origin, and length of marked and unmarked juvenile Chinook salmon in the surface waters of greater Puget Sound. Transactions of the American Fisheries Society 140:170-189.
Ruckelshaus, M. H., K. P. Currens, W. H. Graeber, R. R. Fuerstenberg, K. Rawson, N. J. Sands, and J. B. Scott. 2006. Independent populations of Chinook salmon in Puget Sound. U. S. Department of Commerce, Seattle.
Strange, J. S. 2010. Upper thermal limits to migration in adult Chinook salmon: evidence from the Klamath River basin. Transactions of the American Fisheries Society 139:1091-1108.
Strange, J. S. 2013. Factors influencing the behavior and duration of residence of adult Chinook salmon in a stratified estuary. Environmental Biology of Fishes 96:225-243.
Weitkamp, L. 2012. Marine distributions of coho and Chinook salmon inferred from coded wire tag recoveries. American Fisheries Society Symposium 76:191-214.
Weitkamp, L. A. 2010. Marine distributions of Chinook salmon from the west coast of North America determined by coded wire tag recoveries. Transactions of the American Fisheries Society 139:147-170.